Movie to introduce our lab 研究室紹介動画
研究室の紹介動画を公開しました!(画像をクリックすると再生を開始します。)

Research 研究内容
植物代謝生理学研究室では, 微細藻類を用いて研究を行っています。 一口に藻類と言っても様々な種類があり、当研究室では以下のラン藻、ハプト藻類・円石藻、ラビリンチュラ類、珪藻、黄金色藻、緑藻を主に用いています。 各研究テーマをクリックすると詳しい説明が展開されます。
研究室の紹介動画を公開しました!(画像をクリックすると再生を開始します。)
植物代謝生理学研究室では, 微細藻類を用いて研究を行っています。 一口に藻類と言っても様々な種類があり、当研究室では以下のラン藻、ハプト藻類・円石藻、ラビリンチュラ類、珪藻、黄金色藻、緑藻を主に用いています。 各研究テーマをクリックすると詳しい説明が展開されます。
ラン藻 Synechocystis sp. PCC 6803
ラン藻は淡水(低浸透圧条件)で生息するものから死海(高塩・高浸透圧条件)で生息しているもの、低温地帯や、高温環境、乾燥条件でも生育、生存できる種が存在していることから、その環境適応能の高さが見て取れます。 我々人間のように移動が可能な生物は、例えば「日差しが強くてちょっと暑いな」と感じたら日陰に移って涼むことが出来ますが、植物のように移動のできない生物は自分にとって快適な場所に移るということができません。 ですから、そのような生物は生育環境が個体にとって不都合になった場合でもその環境で生き残る、もしくはより生活しやすくするための仕組みを発達させてきています。
環境に適応するためには様々なステップが必要です。 その最初の段階は"何かが変わった"と細胞が感じ取ることでしょう。 では、生物はどのようにして細胞外環境の変化を認識し、それを細胞活動に反影させているのでしょうか?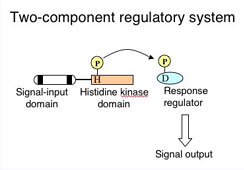 そのような研究は分子生物学的手法が早くから確立された原核生物(特に大腸菌や枯草菌)を材料にして行なわれてきました。 その結果、原核生物では二成分制御系と名付けられた機構が、外部環境の検知、およびその情報伝達に関わっていることがわかってきました。 二成分制御系はバクテリアやカビなのど微生物と植物に保存されており、一般的にヒスチジンキナーゼとレスポンスレギュレーターより構成されるシグナル検知・伝達の機構です。 本研究室では、1996年にゲノムが解読された単細胞性ラン藻Synechocystis sp。 PCC 6803株を用いて光合成生物の環境応答機構の解明を目的に研究を行っています。
そのような研究は分子生物学的手法が早くから確立された原核生物(特に大腸菌や枯草菌)を材料にして行なわれてきました。 その結果、原核生物では二成分制御系と名付けられた機構が、外部環境の検知、およびその情報伝達に関わっていることがわかってきました。 二成分制御系はバクテリアやカビなのど微生物と植物に保存されており、一般的にヒスチジンキナーゼとレスポンスレギュレーターより構成されるシグナル検知・伝達の機構です。 本研究室では、1996年にゲノムが解読された単細胞性ラン藻Synechocystis sp。 PCC 6803株を用いて光合成生物の環境応答機構の解明を目的に研究を行っています。
一つの大きなテーマとして、環境シグナルの検知・伝達系である二成分制御系の解析をしています。 本グループではSynechocystisゲノムに存在する全ての二成分制御系遺伝子について、可能なものについて遺伝子破壊株を構築、解析してきました。 DNAマイクロアレイを用いた網羅的な遺伝子発現解析の結果、いくつかの環境変化に対して応答する二成分制御系を同定しています。
興味深いことに、一つのヒスチジンキナーゼが種々の環境変化に応じて複数のレスポンスレギュレーターにリン酸基を転移する系も発見されています。 現在、その遺伝子産物がどのようにしてそのような制御を行なっているのかについて解析を進めています。
また二成分制御系はある種の藻類の葉緑体ゲノムにも保存されていることが分かっており、その制御系が葉緑体ゲノム中のどの遺伝子の発現に関与しているかに興味をもって研究もしています。
脂肪酸は全ての光合成生物が合成する脂質の1つであり、主に生体膜(細胞膜やチラコイド膜など)の構成成分として知られています。特に炭素骨格中に複数の二重結合をもつ「多価不飽和脂肪酸」は、生体膜の膜流動性に関わる重要な脂肪酸であり、また生物の低温ストレス耐性の獲得に寄与することがわかっています。例えば健康食品として有名なDHAやEPAは、それぞれ炭素数22で二重結合の数が6、炭素数20で二重結合の数が5個の多価不飽和脂肪酸です。
本研究室では、ラン藻Synechocystisをプラットフォームとして、脂質合成に関わる機能未知遺伝子の解析を行っています。脂肪酸種の中には一部の藻類やバクテリアのみでしか検出されていないものがあり、その合成経路や機能を解明するため、ラン藻でそれらの脂肪酸を合成させ、形質転換体の表現型を観察しています。
円石藻 Emilanina huxleyi
植物は、共生により葉緑体(=光合成能力)を獲得した集団であり、系統的に異なる様々な生物の集まりです。そのため、植物の光合成を理解するためには、一つの分類群だけでなく、異なる様々な分類群における研究が必要です。しかし、光合成の研究は緑色植物を中心に進められており、その他の植物や藻類ではまだまだ遅れています。また藻類の中には、緑色植物とはまったく異なる系統に属しており、かつ地球環境に大きな影響を与える重要なグループが存在します。円石藻もその中の1つです。
円石藻は、海洋に生息する単細胞藻類であり、光合成と石灰化により海洋のCO2を固定します。また円石藻は、 “ブルーム(水の華)”と呼ばれる大規模増殖を引き起こします。そのため、地球規模の炭素循環に大きな影響を及ぼす重要な生物であることが知られています。しかしながら、円石藻の光合成(特に炭酸固定)に関する生化学的研究は遅れており、代謝レベルでどのような特徴があるかは明らかになっていません。そこで、私たちの研究室では、円石藻の光合成炭素固定系を、分子レベルで明らかにすることを目的とした研究をおこなっています円石藻の炭酸固定系の解明は、ブルーム形成の要因や、光合成の進化と多様化を考える上で欠かせない、重要な研究です。
 生物による鉱物形成反応をバイオミネラリゼーション(biomineralization)と言います. バイオミネラリゼーションは細菌から植物, 動物にいたるまでさまざまな生物が持っている能力です. 生物によって形成される鉱物は無機的に生じた場合とは結晶の構造が異なり, それにともなって性質も変化することが観察されています. 人の骨や歯はリン酸カルシウム, 貝殻は炭酸カルシウム, イネ科の植物が形成するプラントオパールはケイ酸塩というようにバイオミネラリゼーションによって形成される鉱物の種類もさまざまです. この中でも炭酸カルシウムは海洋生物が形成する鉱物としては最大の割合をしめると推定されています.
生物による鉱物形成反応をバイオミネラリゼーション(biomineralization)と言います. バイオミネラリゼーションは細菌から植物, 動物にいたるまでさまざまな生物が持っている能力です. 生物によって形成される鉱物は無機的に生じた場合とは結晶の構造が異なり, それにともなって性質も変化することが観察されています. 人の骨や歯はリン酸カルシウム, 貝殻は炭酸カルシウム, イネ科の植物が形成するプラントオパールはケイ酸塩というようにバイオミネラリゼーションによって形成される鉱物の種類もさまざまです. この中でも炭酸カルシウムは海洋生物が形成する鉱物としては最大の割合をしめると推定されています.
円石藻はハプト植物門に属する海洋性微細藻類の一種であり, 炭酸カルシウムを主成分とする「殻」を持っています. 私たちの研究室ではおもに円石藻Emiliania huxleyiを使用していますが, この円石藻の「殻」は炭酸カルシウムを主成分とする楕円状の構造物が組み合わさった形をしています. この構造物をココリス(円石)といいます.
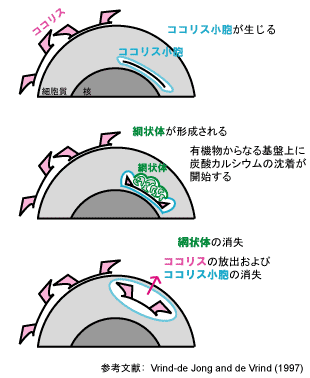
ココリスの形成は細胞内で行われます. 最初に, ココリスの形成の場となる細胞小器官(ココリス小胞) が生じます. この膜構造はゴルジ体由来であると考えられています. ココリス小胞内部には有機物からなる楕円形の基盤が観察されます. 次に, 基盤の辺縁部に炭酸カルシウムの結晶が形成されます. このときココリス小胞に加えて, 網状体とよばれる膜構造が観察されます. 結晶成長が終了した後, 網状体は消失します. 最後に, 完成したココリスは細胞外に放出されます.
ココリスの形成の過程をたどると, ココリス形成には特殊な膜構造の形成や, 大量のカルシウムの輸送など, さまざまな仕組みが必要であることが予想されます. しかし, これらの大半はいまだに明らかになっていません. また, ココリスを溶かすと複雑な構造をもつ酸性多糖が得られることから, 多糖がココリスの形態制御に関わっていることが示唆されています. しかし, この酸性多糖の合成経路もその大半が不明の状態です.
私たちのグループではココリス形成過程に関与する生体高分子の解析を行うことにより, ココリス形成制御機構の解明を試みています.
円石藻は海洋性の単細胞藻類であり、炭酸カルシウムを主成分とする円盤状の特別な構造体(円石)で細胞表面が覆われているのが特徴的です。その中でもEmiliania huxleyiは毎年、衛星観測される程の大規模なブルーム(爆発的な増殖)を生じることで有名です。また、先行研究より生育にセレン(Se)が必須であることが明らかになっています。
セレンは同族の硫黄と物理化学的な性質が非常に似た元素であり、人を含む哺乳類やバクテリアなどの様々な生物で必須微量元素になっています。このSeの必須性は、システインのセレン態であるセレノシステインを酵素の活性中心に特異的に配置した、セレノプロテインと呼ばれるタンパク質の活性維持のためと考えられています。一方で,高濃度に存在すると非常に強い毒性を示すことは生物界共通の現象です。
光合成生物の中で最もSeの研究が活発に行われているのは陸上植物であり、その代謝系も明らかとなっています。しかし陸上植物におけるSeの挙動は、毒性を回避するために無毒な形態で蓄積されることにとどまり、Seの生育に対する必須性やセレノプロテインの存在は認められていません。光合成生物でそれらの報告があるのは一部の藻類に限られており、Seを必須とする光合成生物でSeがどのような機能を果たしているかはほとんど分かっていないのが現状です。
そのため光合成生物におけるSeの生理機能を解明することを目的とし、円石藻Emiliania huxleyiを用いて研究を行っています。さらには、生物の系統分類学上特異な位置にある円石藻のSe代謝機構を解明することによって、生物進化におけるSeの利用戦略の解明に新たな知見を加えることが期待されます。
ハプト藻 Tisochrysis lutea

ハプト藻・円石藻の仲間には「アルケノン」という少し変わった脂質をつくるものがいます。アルケノンは一般的に炭素鎖が37~40程度の長鎖のケトンで、分子内に2~4個のトランス型の不飽和結合を持っています。生体脂質の不飽和結合のほとんどがシス型であることからアルケノンは非常にユニークな脂質であると言えます。アルケノンの不飽和度は合成時の培養温度と強い相関があることから、地質学や有機地球化学の分野では堆積物や地層形成時の温度を推定する古水温計として多用されてきました。しかし、どうしてハプト藻・円石藻がアルケノンを合成するのか、どのように合成・分解されるのかについてはほとんど分かっていません。アルケノンを合成する種はハプト藻や円石藻の中でも限られており、アルケノン合成能はハプト藻・円石藻の進化の過程で比較的新しく獲得されたと考えられています。
私たちはアルケノンを生産するハプト藻Tisochrysis luteaを用いて、重イオンビーム照射による変異育種や遺伝子組換え技術を確立し、生理機能や代謝機構を解明することを目標に研究を行っています。
最近、T. luteaへの重イオンビーム照射により、「フコキサンチン」という光合成補助色素を生合成しなくなった変異株の作出にも成功しました。フコキサンチンは光捕集アンテナの構成要素として光合成において重要な役割を果しているだけでなく、人体への健康効果あることが知られています。一方で、ハプト藻におけるフコキサンチンの生合成経路については未解明なことが多く残されています。私たちが作出した変異株はT. luteaのフコキサンチン生合成経路に新たな知見を与えるものであり、ハプト藻への重イオンビーム照射による変異育種研究は広がりを見せています。